
Glutamate receptors
The most important excitatory neurotransmitter (substance that enables communication between nerve cells) is glutamate. Approximately half of all excitatory synapses in the brain (and there are trillions of them) use this amino acid to transmit information. The fact that you can read this, listen music, and most importantly, that you can remember tomorrow what you read here today, means that there's probably nothing wrong with your glutamate system.
However, there are people with serious disturbances in glutamate receptor function. This can contribute to neuro-psychiatric diseases such as schizophrenia, Alzheimer's dementia or even stroke. That's why we want to know as much as possible about how glutamate system works.
Glutamate acts on the cell membrane structures called glutamate receptors. What we are studying is one group of these receptors that, when activated by glutamate, allow ions to flow into the cells. Therefore, they are called ionotropic receptors. This ionic current then acts like a piece of information, because it tells the cell what to do and how to change its activity, even for very long periods of time. For example, it is believed that these long-term changes in neural activity are a basis for memory storage in the brain.
The ionotropic glutamate receptors we are studying can be subdivided into three groups, based on how they work, what stimulates them and what they are composed of. This latter property is the essential one, because it determines all other receptor characteristics. For example, due to their particular composition, each of the three groups is specifically activated by certain substance, so they are named after them: NMDA, AMPA and kainate receptors.
We are primarily interested in the properties of NMDA receptors. In recent years, our knowledge about the subunits that these receptors are composed of has expanded greatly, so now we can even tell what type of NMDA receptors is present in a particular neuron. It is now known that each receptor is composed of four subunits (Fig. 1). However, these, so called native receptors are not isolated structures in a healthy cell. There are many other proteins attached to the NMDA receptors and for most of these proteins it is still unknown how they influence the receptor activity.

Fig. 1 Structure and subunit composition of NMDA receptors. NMDA receptors consist of four subunits (tetrameric arrangement) - usually comprising two copies each of NR1 and NR2 or NR3 subunits. Every single subunit contains an extracellular N-terminal domain, four membrane domains and an intracellular C-terminal domain. The most protruding part of the N-terminal domain forms various binding sites for different modulatory compounds, such as protons, polyamines (spermine, spermidine), neurosteroids, Zn2+, Cu2+ etc. The binding sites for agonist (glutamate) and coagonist (glycine) are located in the center of the N-terminal domain of NR2 and NR1 subunits, respectively. The transmembrane part of the receptor protein consists of three transmembrane domains (M1, M3, M4) and M2 being a re-entrant loop entering and exiting the membrane from the intracellular side and forming the ion channel pore. The large C-terminal domain mediates interactions with many intracellular proteins, including signaling molecules and proteins involved in protein trafficking, their phosphorylation or dephosphorylation. Interactions of NR1 and NR2 subunits with some of them are shown. To simplify things, only one NR1 and one NR2 subunits are depicted on the right panel.
To simplify things, we occasionally use the cells that normally don't have NMDA receptors, such as human embryonal kidney (HEK) cells. Using a process called transfection, we make these cells create NMDA receptors by putting genes for different subunits in them. In this way, under controlled circumstances, we can study only those recombinant receptors that we want to.
Even more importantly, we don't have to put genes for ordinary NMDA receptor subunits found in nature, but we can play with the genes, change their structure and see what happens with the receptor function. For example, we can take a piece of subunit A and a piece of subunit C, glue them together and create what is called a chimeric NMDA receptor (because of chimera, mythological animal that was part-lion, part-human and part-dragon). Or, we can change just a single amino-acid in the protein subunit structure and see how important it is for a certain aspect of receptor activity (so-called point mutations).
Neurosteroids keep your brain in shape Everybody knows that some hormones in the body have steroidal structure. Steroidal hormones have become notorious because they are occasionally abused by not-so-honest athletes or body-builders. However, even our brains produce steroidal substances that can change the activity of various receptors, including NMDA receptors. In such case, these substances are called neurosteroids.
Recently, in cooperation with dr. Hana Chodounska and her staff (Institute of Organic Chemistry ASCR v.v.i.), we have been testing the effects of two of such endogenous substances, with a similar chemical structure, but with radically different modulatory effects on NMDA receptor activity. One of them is called pregnenolone sulfate (PS) and the other is called pregnanolone sulfate (3α5βS), we use this abbreviation to indicate the positions of hydrogen atoms at positions 3 and 5 in the steroidal ring strucure) (Fig. 2, steroidal structure). To test these substances, we used patch-clamp recording technique. In order to apply agonist, we used the rapid application system.
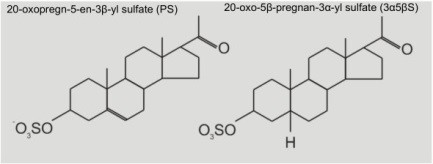
Fig. 2 Structure of pregnenolone sulfate (PS) and pregnanolone sulfate (3α5βS).
An interesting finding was that, for both of these substances, the final effect depends on the mode of application relative to application of glutamate, although in a different manner. Also, we found that the intensity of the effect depends on NMDA receptor subunit composition.
For PS, we have found that when it is pre-applied in the absence of agonist (glutamate or NMDA), the receptor response is potentiated. However, if it is applied simultaneously with the agonist, it still produces potentiation of NMDA receptors that contain subunit NR2A or NR2B, but it also slightly inhibits the activity of NR2C or NR2D containing receptors (Fig. 3). As you can see, glutamate itself produces a control response. The line is below zero level, i.e. the ionic current is negative to indicate that positive ions flow through the NMDA receptor channel into the cell. Then, when PS is pre-applied, the current becomes larger. The higher the concentration of PS, the larger is the response. With the highest PS concentration we used (300 µM), the responses are potentiated 4 to 5 times relative to control.

Fig. 3 The effect of PS on NMDA receptors made of different subunits. Upper panel: When only glutamate is applied (indicated by black bar) to NR2A-containing receptors, the cell responds by control response. Then, when PS is pre-applied before glutamate, the response is potentiated approx. 3 times. However, when PS is applied both before and during glutamate application, the response obtained is somewhere between control and pre-application responses. This means that PS also slightly inhibits NMDA receptors, but only when they are activated by glutamate. Finally, to show that these results are really a consequence of PS and not some accidental change in the cell, another glutamate-only response is recorded and it looks the same as the initial one. Lower panel: The mode of application of PS and agonist is the same as above, but the receptors are different: now they contain NR2C subunit. As can be seen, this type of NMDA receptor is only slightly potentiated by PS pre-application, but it is strongly inhibited when PS is applied simultaneously with glutamate.
For 3α5ßS, we have found that it inhibits NMDA receptors, regardless of their composition, but only when it is applied simultaneously with the agonist, as shown in Fig. 4. However, our interesting finding is that 3α5βS cannot bind to the NMDA receptor when applied in the absence of the agonist. This is a particularly interesting property that 3α5βS seems to share with a group of drugs called open channel blockers. As previously said, NMDA receptors actually contain a channel through which the ions flow into the cell. In the absence of the agonist, the receptor is inactive and the channel is closed. There are substances that can get stuck in the channel. In this way, they block the flow of ions and, therefore, inhibit the NMDA receptor activity. Since the binding site for such substances is within the channel, they cannot manifest their effect unless the channel is open, i.e. the effect can be seen only if NMDA receptor is „used“ by the agonist. For this reason, these blocking substances are called use-dependent blockers or open-channel blockers.

Fig. 4 Left: When only glutamate is applied (indicated by white bar above the recording), the cell responds by inward current. Then, when 3α5βS is simultaneously applied in high concentration (indicated by black bar above), the response is immediately and almost completely inhibited, i.e. the current precipitously falls back to zero. Then, after cessation of 3α5βS application, the current very slowly recovers back to control level. Middle: when 3α5βS is applied prior to glutamate, no change occurs, i.e. this response looks exactly the same as the control. This means that 3α5βS can not inhibit receptors that are not activated („used“) by glutamate. Right: Overlapped boxed regions a and b show differences in responses after 3α5βS application, depending on whether NMDA receptors where activated or not.
However, our additional finding is that the binding site for 3α5βS doesn’t seem to be within the channel, because its effect does not depend on membrane potential, as is the case for open-channel blockers.
We also tested the effects of 3α5βS on single NMDA receptor channels. In general, any substance can change one of the three properties of single channels (or a combination of these): the frequency of channel opening, channel conductance and/or duration of time during which the channel remains open (called open time). We have shown that 3α5βS inhibits the overall NMDA receptor activity by primarily reducing the frequency of channel opening, but also by slightly shortening the open time.